2023.1.11(综述)LncRNA的定义、功能、挑战和建议


这篇文章讨论了lncRNA的定义、命名、保守、表达、表型可见性、功能分析和分子机制,包括lncRNA与染色质结构、表观遗传过程、增强子功能和生物分子凝聚,以及lncRNA在核外的作用。
lncRNA包括 Pol I,Pol II 和 Pol III 转录的RNA,以及来自加工过的内含子的RNA。
大多数lncRNA比蛋白质编码序列进化得更快。
很多lncRNA和染色质修饰复合物相关。
lncRNA的作用:在细胞核中,空间上调控发育过程中的基因表达;在细胞质中,参与调控、代谢和信号转导的调控。
lncRNA通常具有一个模块化结构并且富集重复序列,这往往和它们对应的功能相关。
G值悖论:生物的 “基因数目即G值” 与生物的复杂度并不正相关。
C值悖论:生物的 “基因组大小即C值“ 与生物的复杂度并不正相关。
秀丽隐杆线虫由 ~1,000 个体细胞组成,而人类由 ~30×10^12 个体细胞组成,两者都有 ~20,000个蛋白质编码基因。
非编码DNA的范围,非编码RNA的转录,都随着发育复杂性的增加而增加。
最开始,这些unusual RNA被认为是转录噪音。后来,越来越多的人报道lncRNA的动态表达和生物学功能。随着技术发展,人们能够鉴定并区分这些RNA。有小部分lncRNA虽然有可靠的注释但是很少有机制信息。
这篇文章的作者认为表达lncRNA的位点应该被识别为真正的基因。
lncRNA的定义:
含有超过200个核苷酸的非编码转录本。包括了5SRNAs、tRNAs、snRNAs和snoRNAs,以及miRNAs、siRNAs和piRNAs。
这个定义排除了其它一些短RNA,如:
- snaRs,~80-120nt,和核因子90相关;
- Y RNAs,~100 nt,作为核糖核蛋白(RNP)复合物的支架;
- vault RNAs,88-140nt,参与将细胞外刺激转移到细胞内信号;
- 其它的由转录后加工产生的启动子相关的RNA和非典型小RNA;
- 其它接近200nt的非编码RNA,如7SK(脊椎动物中 ~330nt,控制转录平衡和终止,包括增强子);7SL(~300nt,信号识别粒子的一个组成部分,将蛋白质靶向到细胞膜上);Alu进化祖先等 这篇文章的作者把非编码RNA非为三类: 第一类是:小RNA(小于50nt) 第二类是:RNA聚合酶III转录本(如tRNAs、5S rRNA、7SK、7SL、和Alu、vault和Y RNA),植物中的Pov V转录本和小的RNA聚合酶II转录本如大多数snRNA和内含子衍生的snoRNA(~50-500nt) 第三类是:lncRNA(超过500nt),它们主要是Pol II产生的
许多lncRNA因为被剪切和多聚腺苷化而被描述成“mRNA-like”。然而,其它lncRNA不是聚腺苷化或具有7-甲基鸟苷帽,它们是从Pol I或Pol III启动子表达或者是从前体加工而来,包含内含子和重复元件,导致未知功能的转录本。
对于蛋白质编码基因,lncRNA可以是 基因间,反义,或者是内含子。它们也来源于“假基因”,这种基因通常存在于后生动物的基因组中,小鼠基因组中发现了超过10,000个假基因,人类基因组中发现了将近15,000个,其中一些已被证明具有功能。
lncRNA还包括由编码和非编码转录本反向剪切产生的环状RNA,已被证明也有功能,以及反式调控RNA,通常作为mRNA的3'端非翻译区。
根据lncRNA相对于蛋白质编码基因的基因组位置和方向对其进行命名和分类。
在人类和黑腹果蝇中,许多蛋白质编码基因都有5'端外显子,在早期胚胎发育中整合到mRNA中。并位于通常的第一个外显子的上游成百上千个碱基,绕过了在过渡区域的许多其它基因。事实上,任何碱基都可能是外显子,内含子,或“基因间”,这取决于细胞在其发育轨迹或生理状态中的任何时刻的转录输出。因此,除非lncRNA和蛋白质编码基因是反义的,否则我们建议根据lncRNA本身的特征或功能来命名它们,如:XIST,AIRN(反义链IGF2R非编码蛋白RNA),HOTAIR(HOX反义链基因间RNA),Gomafu,COOLAIR(指植物的春化作用),APOLO(生长素调节启动子环)。为了便于回忆,最好有完整的外显子-内含子结构和基因组坐标,如果没有生物学背景,我们建议根据GENCODE系统来命名lncRNA。
非编码RNA的广泛功能排除了直接分类为特定的RNA类别,有些在局部作用,有些在远处作用,或者两者兼有。非编码RNA有各种形状和大小,而且面积很大,覆盖了大部分的基因组和过多的功能。一些RNA有双重作用,比如同时作为编码RNA和调控RNA,并且还有一些,可能是许多的胞质lncRNA编码小肽。蛋白编码位点也通过选择性剪切表达lncRNA,令人惊讶的是,约17%的人类蛋白编码位点产生的主要转录本都是非编码的。事实上,lncRNA基因和mRNA基因都可以产生转录本,并在不同程度的加工后发挥作用。未剪接的转录本、剪接的转录本、环状RNA、内含子RNA以及由它们产生的稳定的小RNA都可以具有功能。任何RNA都可以是调节性的,任何位点都可以编码蛋白质编码RNA和调节性RNA。
已经有超过10w个人类lncRNA被记录下来,其中许多是灵长类动物谱系特有的。现在有数十万个已编目的lncRNA和数十个数据库。在过去的十年里,已经有约5w篇论文以“长链非编码RNA”为一个关键术语发表,并且有超过2k篇论文报告了有效的lncRNA功能,尽管它们中的大多数都没有在后续得到任何详细的追踪。
接下来,作者主要关注来源于Pol II初级转录单位的lncRNA,而不是来源于Pol I或Pol III启动子表达或经内含子处理的或通过反向剪切形成的其它非编码RNA(需要注意的是,内含子在哺乳动物和其它生物中都构成了非编码RNA的主要部分)。
lncRNA的保守性:
大多数lncRNA在物种间的保守性低于编码蛋白质组的mRNA序列。最开始,大多数哺乳动物基因组(包含大多数lncRNA位点)被认为是中性进化。利用人类和小鼠基因组之间分化率的标准(来自转座子的常见的ancient repeats),假设这些序列是无功能的,并代表了在祖先中的原始分布。然而,越来越多的证据表明转座因子被广泛地团结协作,作为基因表达和结构的功能元件,形成启动子,调节性网络,蛋白质编码基因和lncRNA中的外显子和splice junctions,因此不能作为中性进化的指标。
调节性序列,包括启动子和lncRNA,被认为是快速进化的,因为有比蛋白质编码序列更宽松的结构-功能约束,这是由于在适应性辐射中的正向选择。许多lncRNA都是细胞谱系特异性的,事实上,考虑到它们与发育增强子的关联,lncRNA的补体和序列变化可能是生物多样性的一个主要因素。
位点表达lncRNA展示出蛋白质编码基因的许多特征,包括启动子,多个外显子,选择性剪切,染色质特征,形态因子和传统转录因子的调控,癌症和其它疾病的表达改变,以及一系列类似于mRNA的半衰期。
lncRNA的启动子表现出与蛋白质编码基因相似的保守水平。lncRNA还具有保守的外显子结构,splice junctions和序列补丁,尽管序列进化快速,但它们仍然保留了同源功能。事实上,低序列保守性可能会产生误导。
lncRNA端粒酶RNA模板成分(TERC)是端粒维护所必需的,这是一种重要的细胞功能,在大小和序列上差异很大,但从酵母到哺乳动物都具有保守的拓扑结构,尽管有一些变化,但是保守的催化核心是不变的。果蝇的X染色体剂量补偿,需要通过lncRNA roX1和roX2与特定伴侣蛋白(雄性致死2,MSL2)的内在无序区域(IDR)相互作用进行相分离,形成一个核结构域。哺乳动物同源物MSL2的IDR取代黑腹果蝇蛋白和roX2的表达足以在哺乳动物细胞中形成异位X染色体剂量补偿,表明roX-MSL2 IDR相互作用是X染色体区段化的主要决定因素,并且这种相互作用在巨大的进化距离上被保留。在胎盘哺乳动物中,XIST也参与调节X染色体的剂量补偿过程,其功能包括排斥常染色质因子,搭建新的异染色质因子和重组染色体结构。
表达
虽然也有例外(如MALAT1,也称为NEAT2,它是脊椎动物细胞中最丰富的Pol II转录本之一,以及NEAT1),lncRNA通常比mRNA表现出更多的有限表达模式,并且通常具有高度的细胞特异性,这与在细胞状态和发育轨迹的定义中的作用一致。lncRNA也有特定的亚细胞位置,通常是核,也有一部分是在细胞质。HOX基因的差异表达在表面上相似的皮肤细胞的不同区域中。lncRNA的表达在大脑的不同区域和不同的发展阶段。lncRNA也在哺乳动物干细胞,肌肉,乳腺,免疫和神经细胞的分化过程中动态表达。其中,在发育过程中从广泛表达和保守的lncRNA向越来越多的谱系特异性和器官特异性lncRNA转变。
lncRNA的表达也会受到环境因素的强烈影响,这一特征在植物中尤为突出,包括动物中的一系列应激反应和癌症中的耐药性。
lncRNA在不同发育阶段的不同细胞中的限制性表达及其通常拷贝数通常较低,导致了lncRNA在bulk tissue RNAseq中表现稀疏特征,而许多lncRNA在特定细胞中相对容易检测。lncRNA的取样不足正在被靶向捕获,先进的成像,空间转录组,单细胞测序等修正,目前已有约2w个人类lncRNA位点被GENCODE鉴定出来,3w个被FANTOM组织鉴定出来,真实的有可能至少要多一个数量级。
由于转录起始和终止位点,表达水平和剪切的高度复杂性和变化,对转录组的全面表征是极具挑战性的。最近的一项研究表明,lncRNA的低表达通过确保其调控靶点的特异性,对其功能作用至关重要,这表明低丰度水平可能是lncRNA如何工作的一个基本特征。
表型可见性
与miRNA一样,大多数lncRNA还没有在基因筛选中被识别出来,这主要有两个原因:首先,大多数基因筛选都集中在蛋白质编码突变上,这些突变往往有严重的后果,相比之下,调节性突变经常只有微小的后果。其次,很难在非编码序列中发生的许多变异中识别出因果突变。事实上,大多数影响人类数量性状和复杂疾病的变异发生在非编码区,在那里有很多表达lncRNA的基因。
通过基因鉴定的lncRNA也有例外,特别是参与雄性果蝇X染色体激活的roX1和roX2的RNA,哺乳动物亲本印迹小鼠H19,Airn和Kcnq1ot1的RNA。在拟南芥中,非编码内含子单核苷酸多态性对于开花时间的适应非常重要,改变了lncRNA COOLAIR的剪切。
许多lncRNA和诱发肿瘤和肿瘤进展相关,通过改变致癌基因或肿瘤抑制因子的表达或突变。其它lncRNA也参与了人类遗传病,包括 DiGeorge syndrome,以及其它神经发育和颅面缺陷。苯丙酮尿症,也是人类遗传病,主要是由苯丙氨酸羟化酶突变引起,也由lncRNA突变引起。
分析lncRNA生物学功能的一种途径是沉默或删除,或者是不太常见的异位表达。
在7个人类细胞系中,超过1.6w个lncRNA被转录抑制,确定了近500个正常细胞增殖所需的lncRNA,其中89%仅在一种细胞类型中表达。
(1)小鼠长链非编码RNA基因突变的可见表型
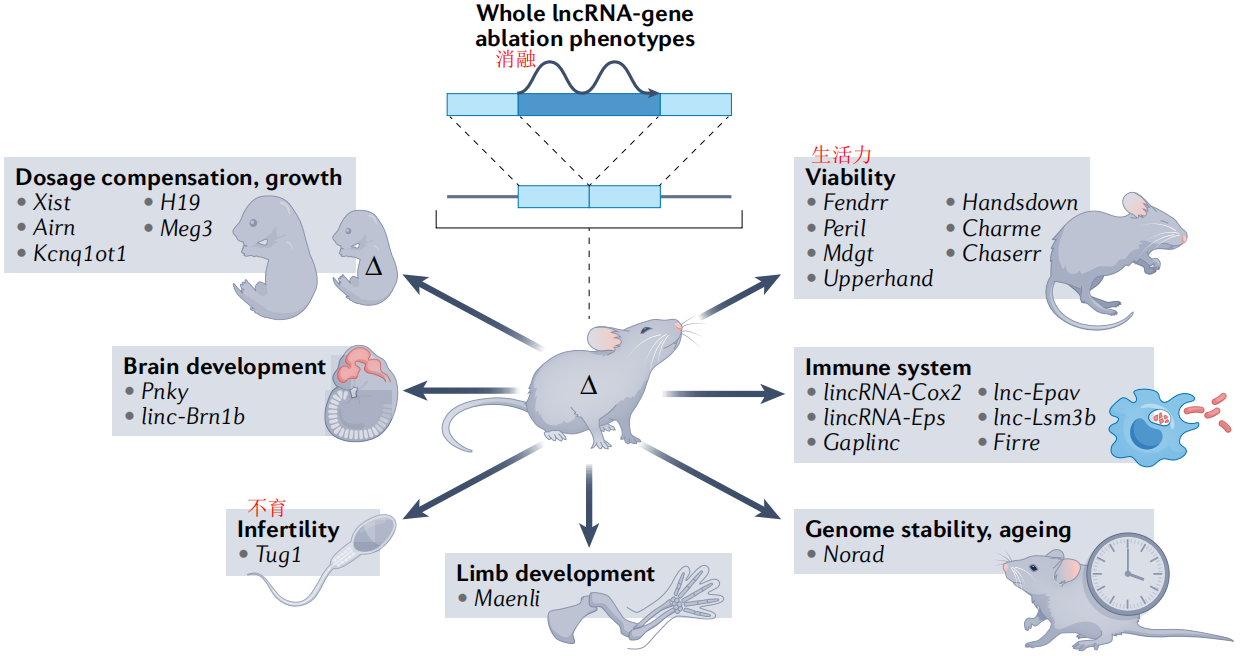
脑细胞质RNA1(BC1)是一种高表达的脑lncRNA,在小鼠中行为改变上看起来无害,但在野外中却是致命的。因此,广泛的表型分析很重要,特别是对于认知功能,类器官模型可能有助于在体外识别表型。
lncRNA的功能注释也可以通过分子表型来进行。对CRISPR-Cas13介导的400多个lncRNA缺失后的表达模式,lncRNA染色质相互作用和其它分子指标的分析表明,lncRNA调控了许多参与发育,细胞周期和细胞黏附等过程的基因。
lncRNA的生物学功能
有例子表明RNA通过RNA-RNA,RNA-DNA和RNA-protein的相互作用,几乎参与了基因组组织,细胞结构和基因表达的所有水平,通常涉及重复元件,包括3'端非翻译区的SINE。这些相互作用包括染色质结构和转录调控,剪切,蛋白翻译和定位,以及其它形式的RNA过程,编辑,定位和稳定性。
许多lncRNA参与动物和植物的细胞分化和发育调控,也参与生理学过程,比如p53介导的DNA损伤应答,免疫细胞中的VDJ重排和种类转换重组,细胞因子表达,内毒素性休克,炎症和神经性疼痛,胆固醇的生物合成和体内平衡,生长激素和催乳素的产生,葡萄糖代谢,细胞信号转导和转运途径,突触功能,以及植物对各种生物和非生物胁迫的响应。lncRNA和细胞膜和核酶之间也有一种新的联系。
目前发表的文献主要集中在:lncRNA在分化和发育中的普遍性和重要性,lncRNA和染色质修饰蛋白的关联,发育性增强子中lncRNA的表达,以及RNA核相分离凝聚物的形成。
染色质结构的控制
在植物中,已知DNA甲基化是由小的非编码RNA指导的。在真菌和动物中,异染色质形成和表观基因沉默都需要RNAi途径。哺乳动物的从头DNA甲基转移酶3A(DNMT3A)和DNMT3B以高亲和力结合siRNA,而不是维持DNA甲基转移酶DNMT1。
DNMT1可以恢复半甲基化CpG的甲基化。
三胸复合物(可使H3K4甲基化)。
先锋转录因子,如NANOG、OCT4(也称为asPOU5F1)。
锌指转录因子的结合位点选择CTCF。
先锋转录因子CBP也能结合RNA,包括那些从增强子中转录出来的RNA,以刺激组蛋白乙酰化,从而刺激转录。一些转录因子(OCT4、NANOG、SOX2和SOX9)也受到lncRNA的调控,包括假基因衍生的lncRNA。
增强子作用
增强子是非编码基因组位点,控制发育过程中的时空表达。在哺乳动物基因组中有~40万个(±10万)个增强子。有时也被归类为 super-enhancers 和 enhancer jungles。
增强子表达双向转录的启动子相关的短RNA。
lncRNA结构和功能的关系
lncRNA的大小一般从大约1 kb到超过100 kb不等,并且都有一个模块结构。选择性剪切已经被证明可以改变lncRNA的功能。
化学探测结果表明,包括Xist在内的lncRNA形成了复杂的多结构域结构。在Xist中,最高度保守的序列是它的重复序列。
(2)长链非编码RNA在核组织中的角色
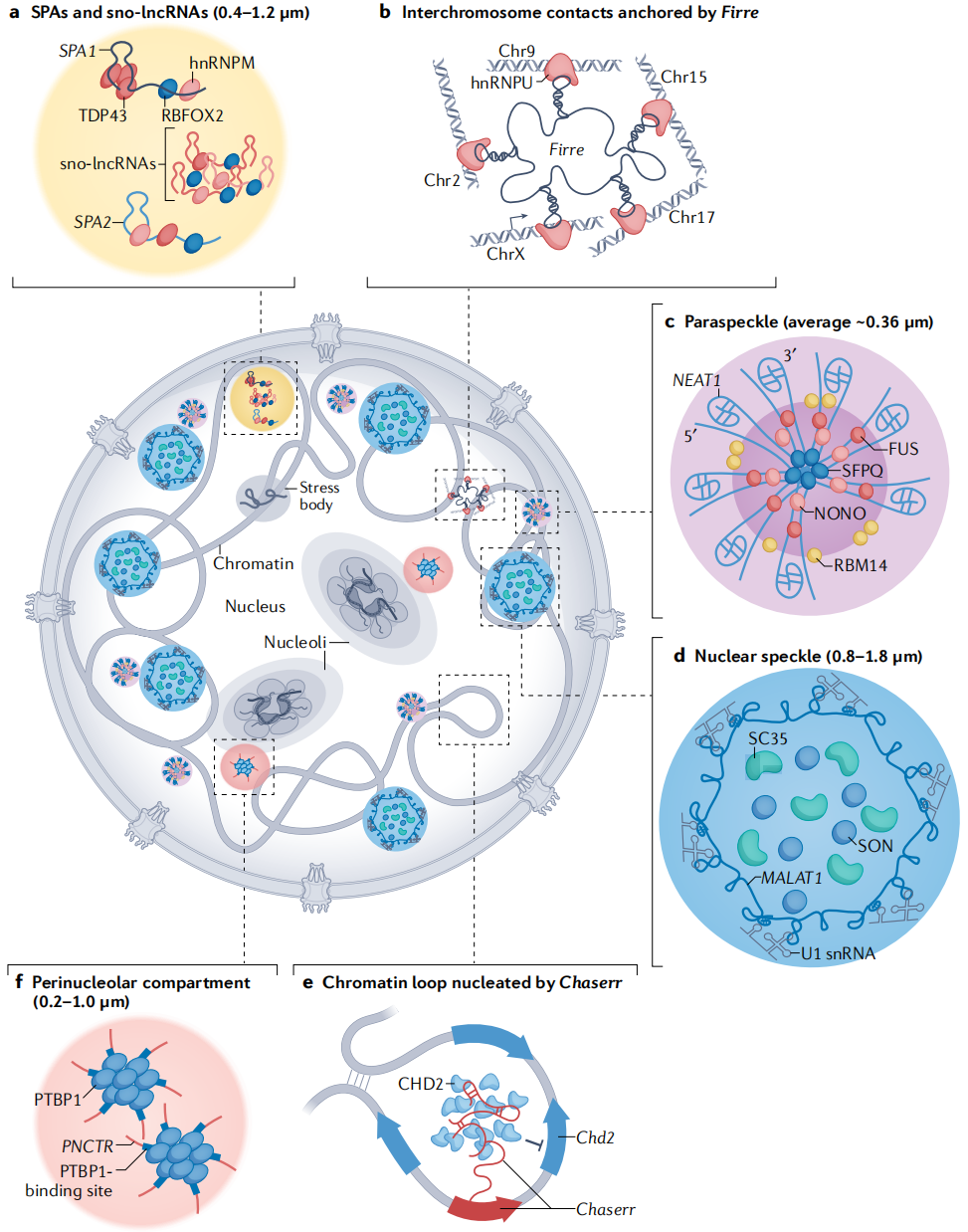
(3)长链非编码RNA的模块化结构
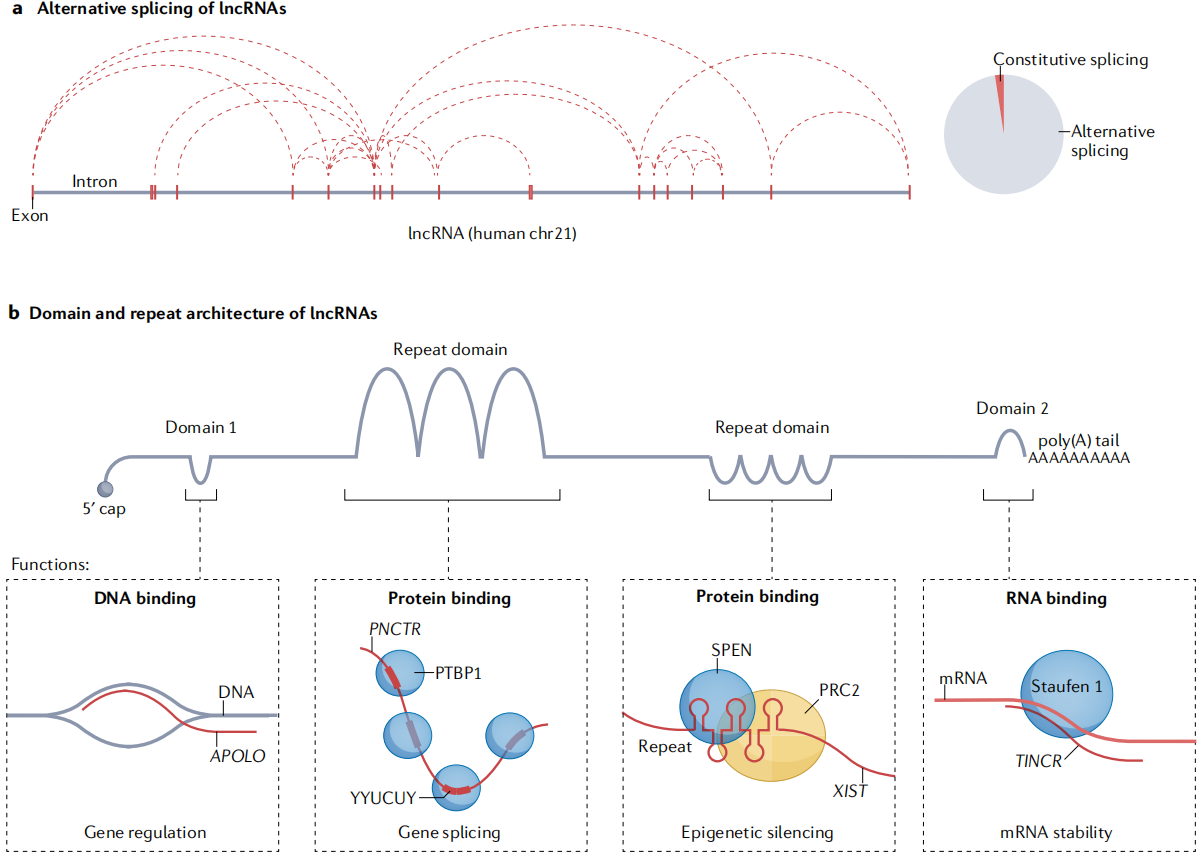
lncRNA作用的分子机制尚不清楚。在大多数特征明确的RNA调控例子中,如RNAi,snoRNA,CRISPR和端粒酶,RNA作为向导,靶向效应蛋白复合物到互补RNA或DNA序列。有证据表明,lncRNA形成三联体是一种普遍存在的现象。其他的,特别是反义lncRNA,似乎通过RNA-DNA杂交形成发挥作用。
本文系外文翻译,前往查看
如有侵权,请联系?cloudcommunity@tencent.com?删除。
本文系外文翻译,前往查看
如有侵权,请联系?cloudcommunity@tencent.com 删除。